文丨粉墨叔叔
前言卵子发育受到从无脊椎动物到哺乳动物等生物体的多种环境刺激的调节。卵子发生受营养可用性的影响很大,因为卵母细胞需要大量的能量来确保成熟卵子的产生。特别是,昆虫卵子生成通常是营养依赖性的,当没有足够的营养时受到严重抑制。
昆虫卵子发育也受到加速产卵的交配刺激的影响。人们认为昆虫卵子发生的营养和交配依赖性是一种适应性现象,因为雌性昆虫尽管体型很小,但交配后通常需要产下大量的大卵。
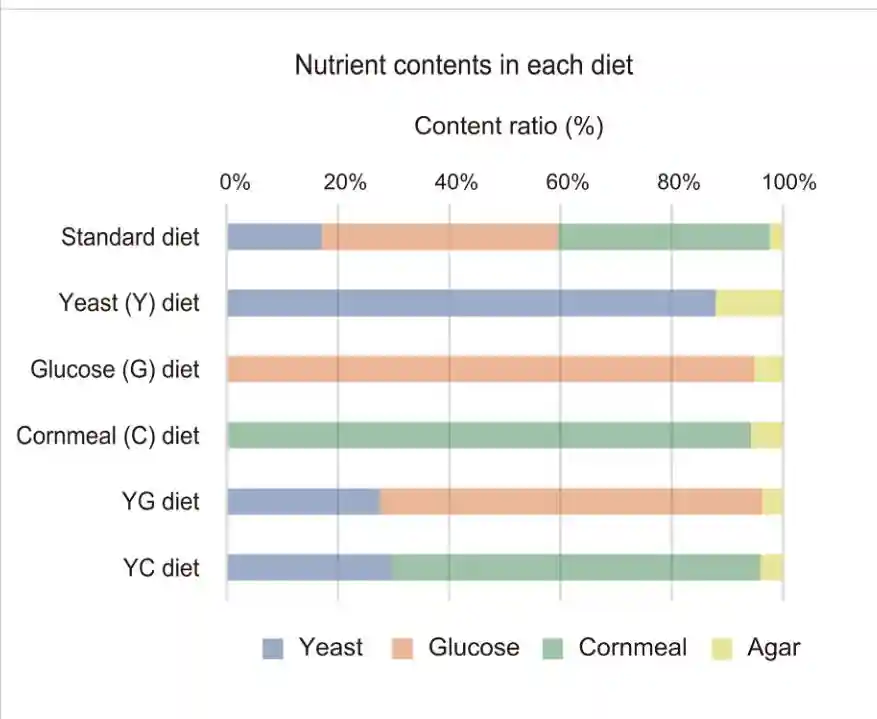
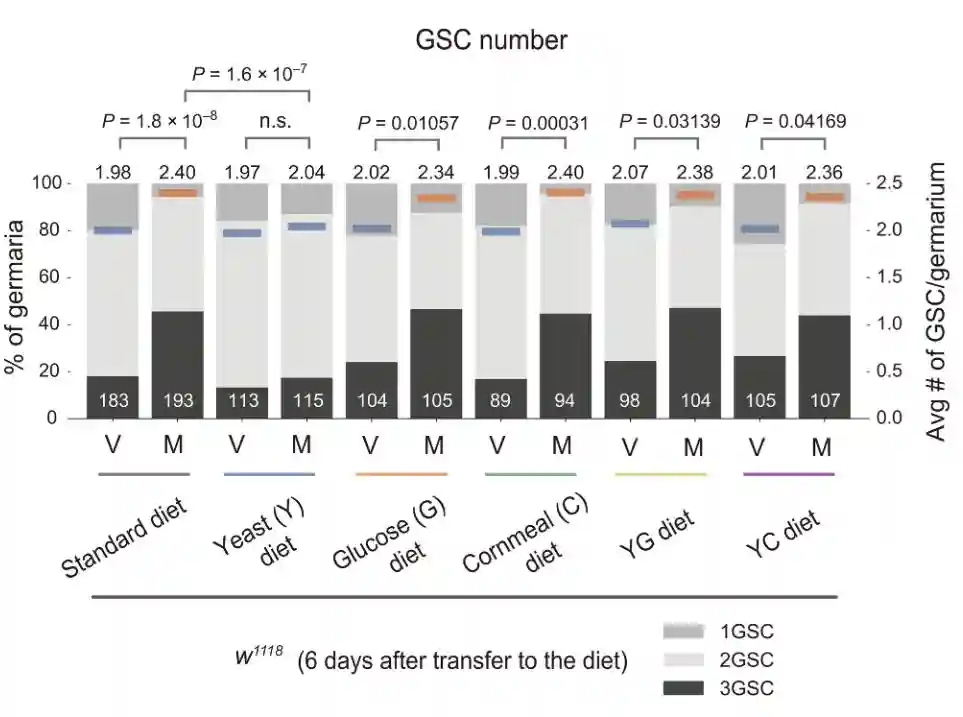
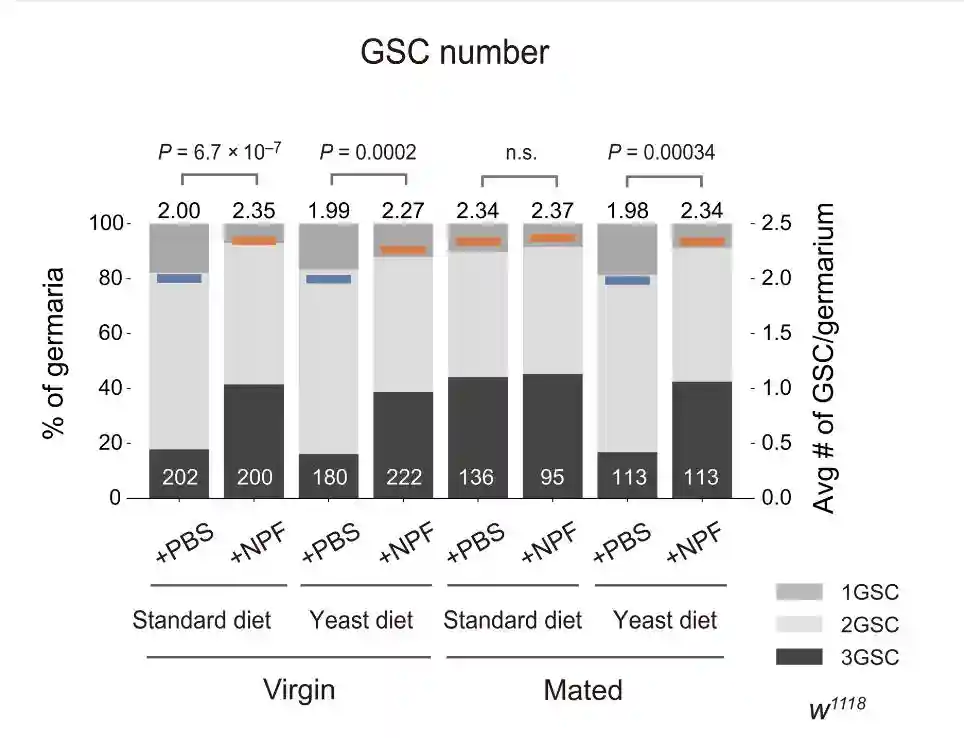
为了研究营养状况对交配诱导的GSC增加的影响,首先比较不同营养成分的苍蝇喂养日粮中交配前后的GSC数量。
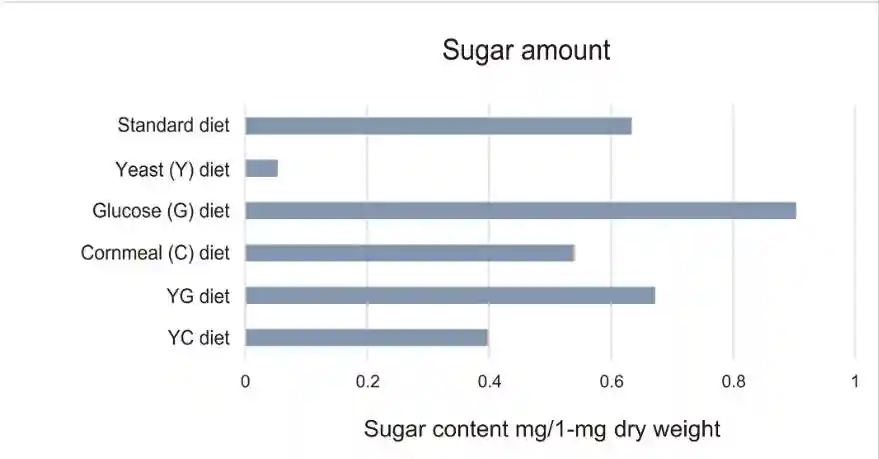
饮食的标准是干酵母、玉米面、葡萄糖和琼脂的混合物。除了标准饮食外,另外还准备了五种替代饮食:干酵母加琼脂,葡萄糖加琼脂,玉米面加琼脂,干酵母和葡萄糖加琼脂。
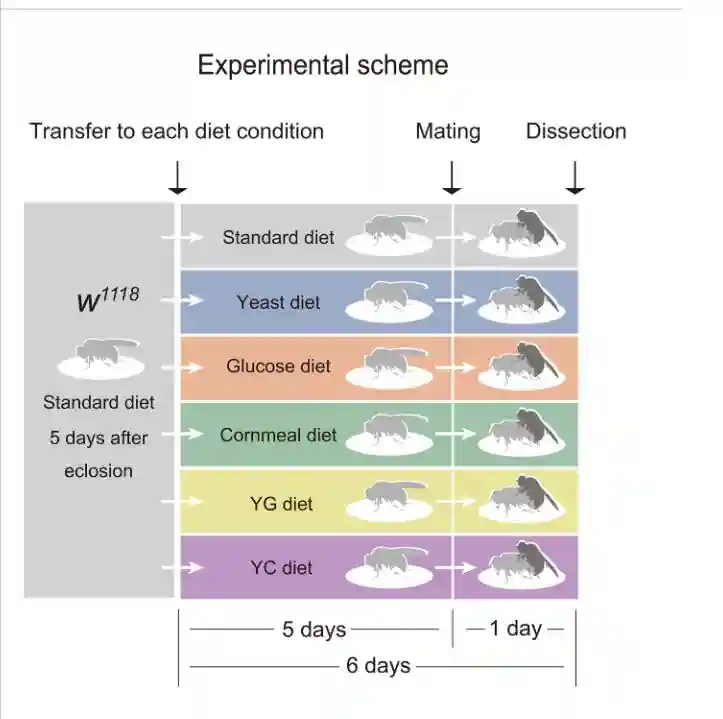
以标准饮食饲养野生型苍蝇,从鸡蛋到年轻人。在封闭后5天收集的年轻雌性被转移到装有标准饮食和五种替代饮食的小瓶中。转移五天后,其中一些苍蝇被允许与雄配1天,其他雌性苍蝇在没有雄性的情况下连续饲养1天。
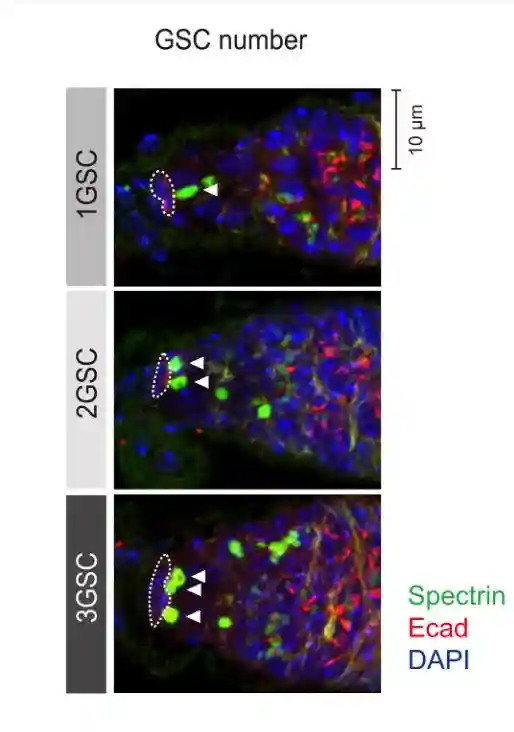
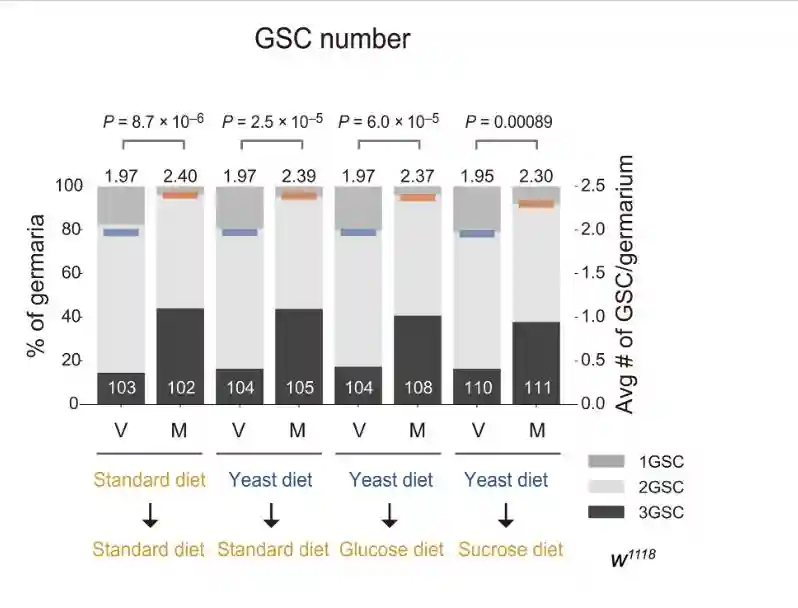
交配诱导了标准饮食饲养的雌性以及葡萄糖、玉米面、YG和YC饮食的雌性GSC增加。相比之下,交配诱导的GSC增加在酵母饮食中饲养的雌性中受损。
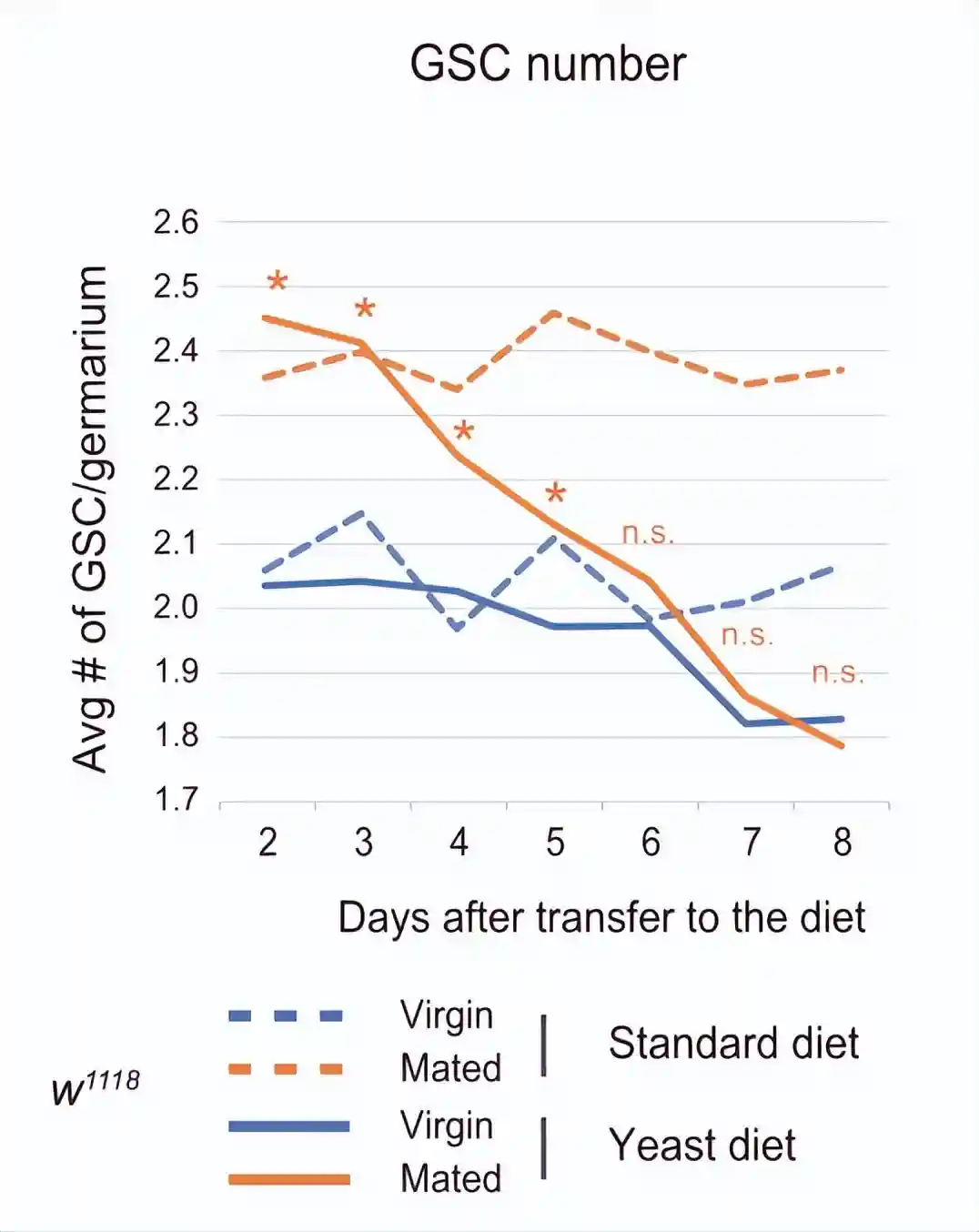
检查转移到酵母饮食后多少天,出现了交配诱导的GSC增加的损害。年轻的雌性以标准饮食或酵母饮食饲养1至7天。每个实验批次的雌性与或不与雄性一起饲养1天。
在标准饮食中,在任何检查的时间点观察到交配诱导的GSC增加。相反,在酵母饮食中,在转移后的前5天而不是6天内观察到交配诱导的GSC增加。
为了排除酵母饮食中的雌性不能与雄性有效交配的可能性,使用dj-GFP雄性,其中精子用绿色荧光可视化,并检查精子是否成功转移到酵母饮食中的雌性身上。用酵母饮食饲养野生型雌性5天。
将雌性与dj-GFP雄性放置1天,超过90%的雌性腹部携带绿色荧光精子。该交配频率与以标准饮食饲养6天的雌性没有差异。结果表明酵母日粮中交配诱导的GSC增加的损害不是由于交配失败。
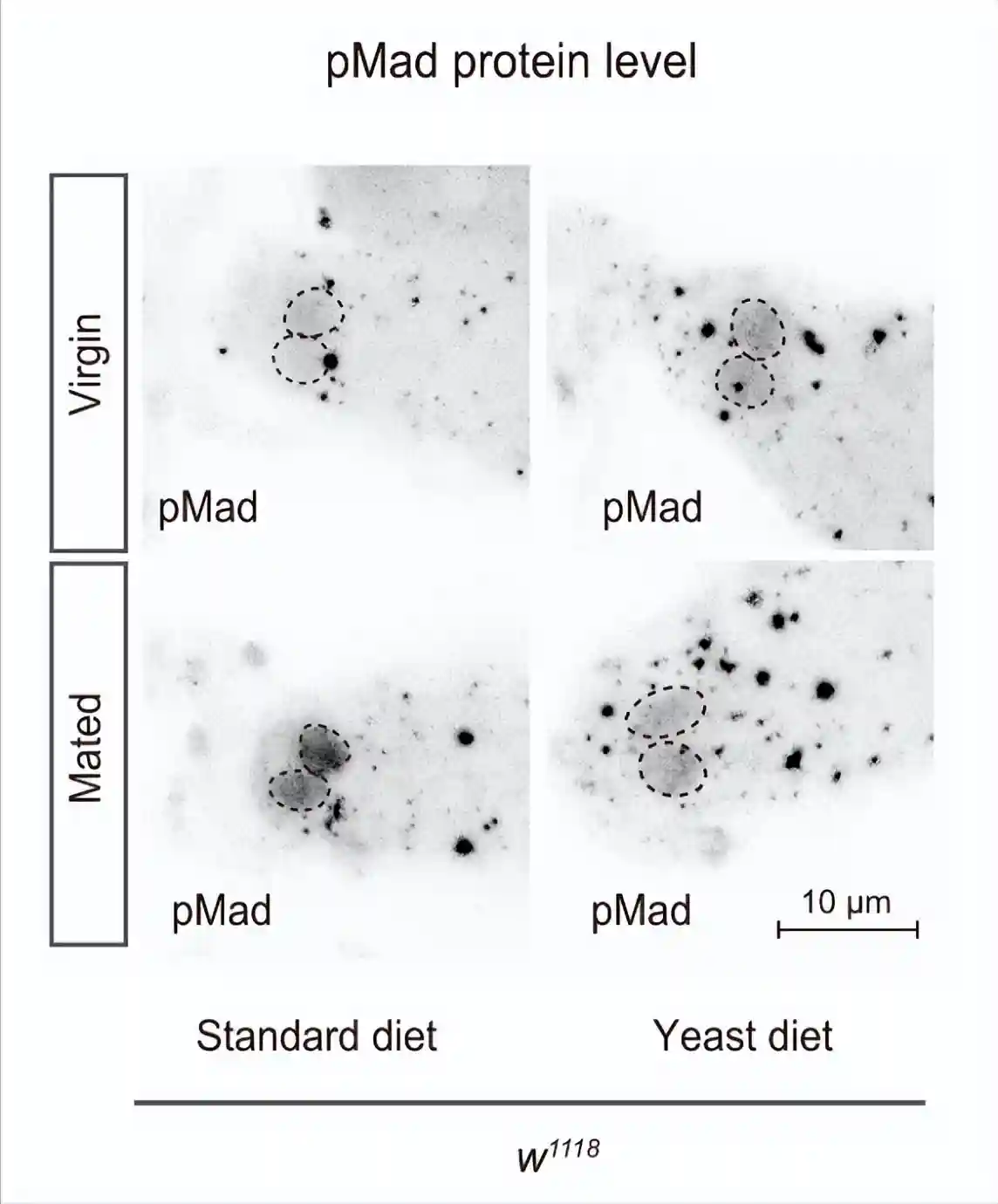
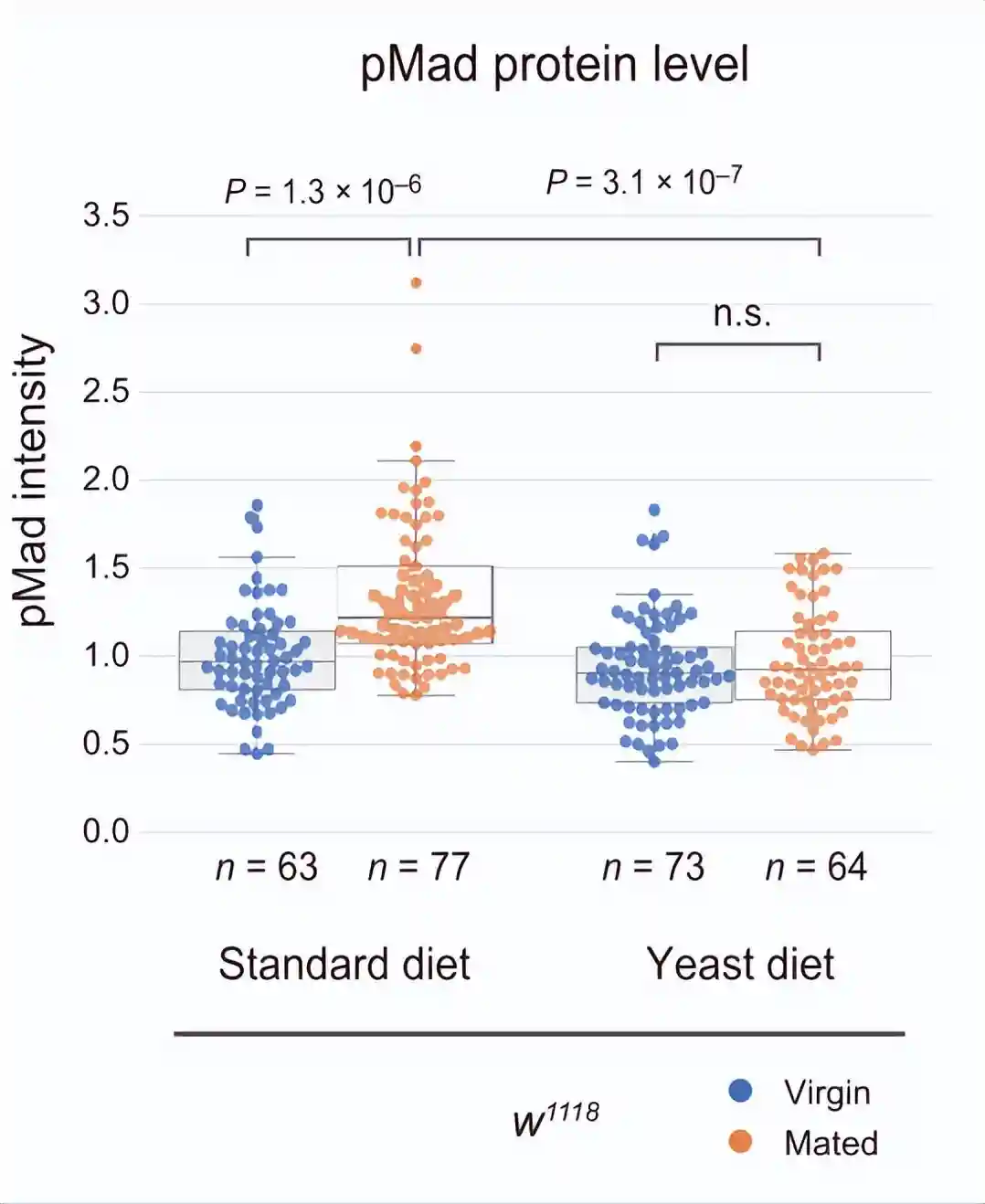
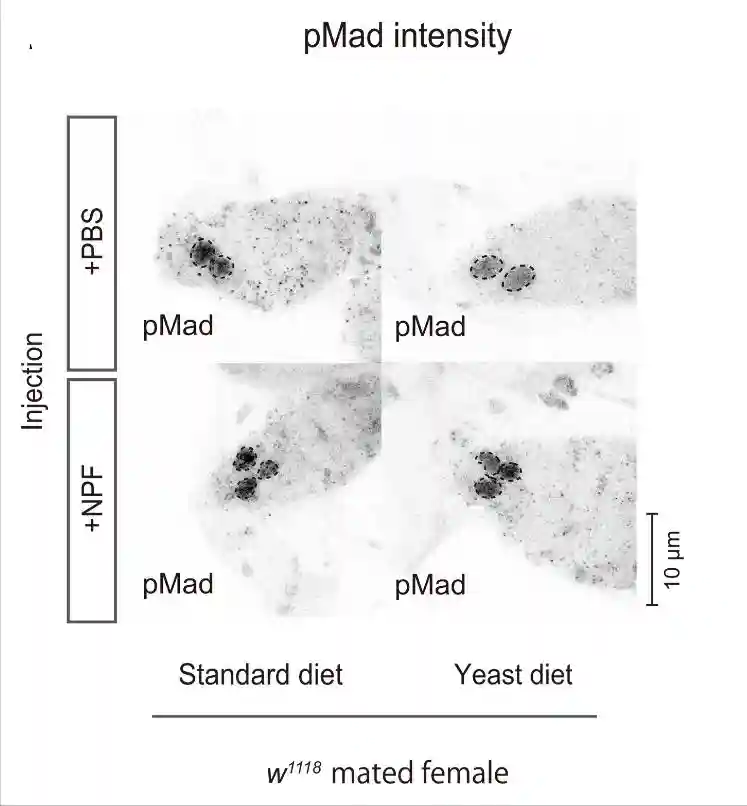
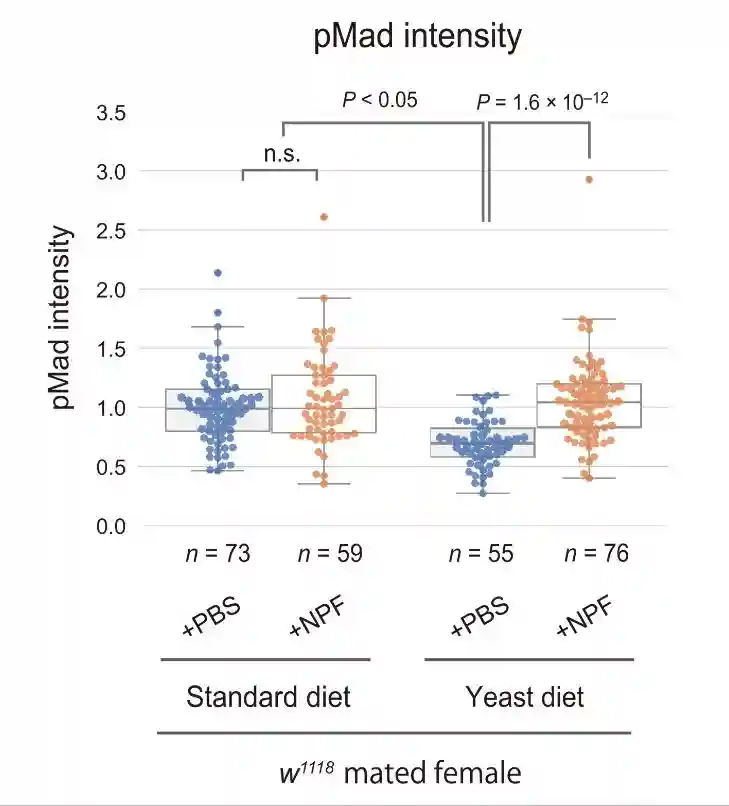
测量GSC中磷酸化母亲相对于dpp的量,当GSC增殖和维持所需的生态位信号十五瘫痪从帽细胞传递到GSC时,它促进Mad的磷酸化以维持GSC的干性。与雌性相比,GSCs中的pMad强度在交配的雌性中升高。
酵母饮食抑制了交配诱导的pMad升高,而标准饮食和酵母饮食中雌性GSC的pMad水平保持不变。这些结果表明,交配诱导的果蝇在酵母饮食中的GSC增加的损害是由于交配诱导的GSC中pMad升高的损害。
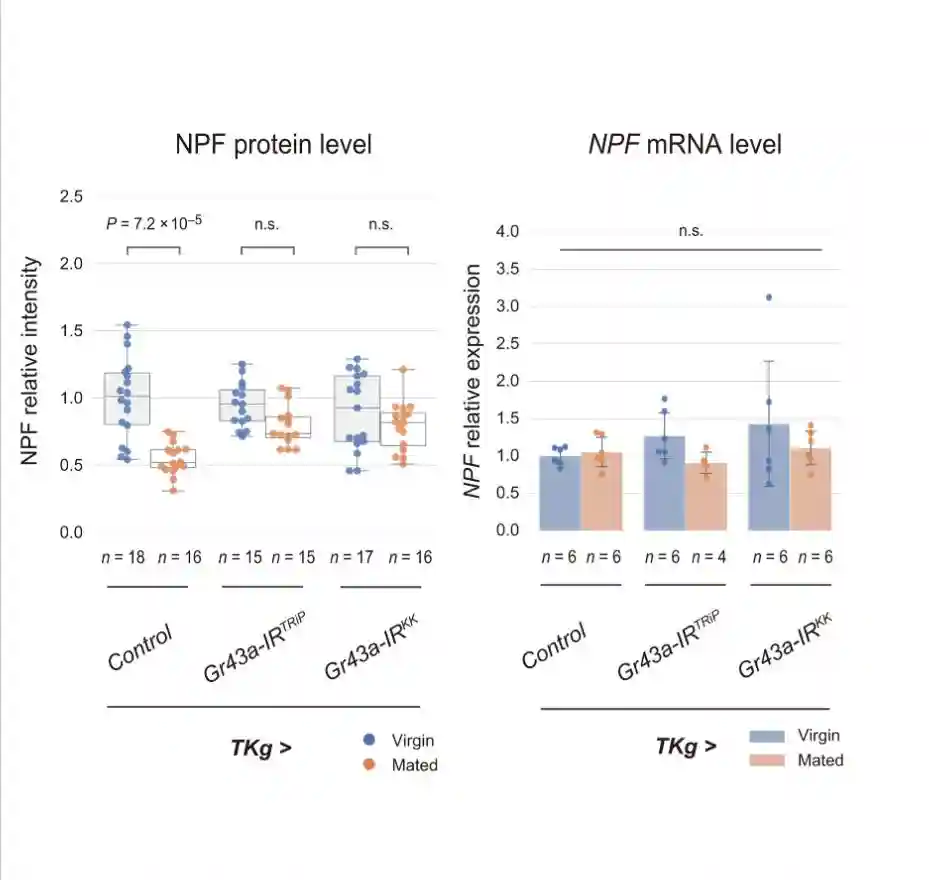
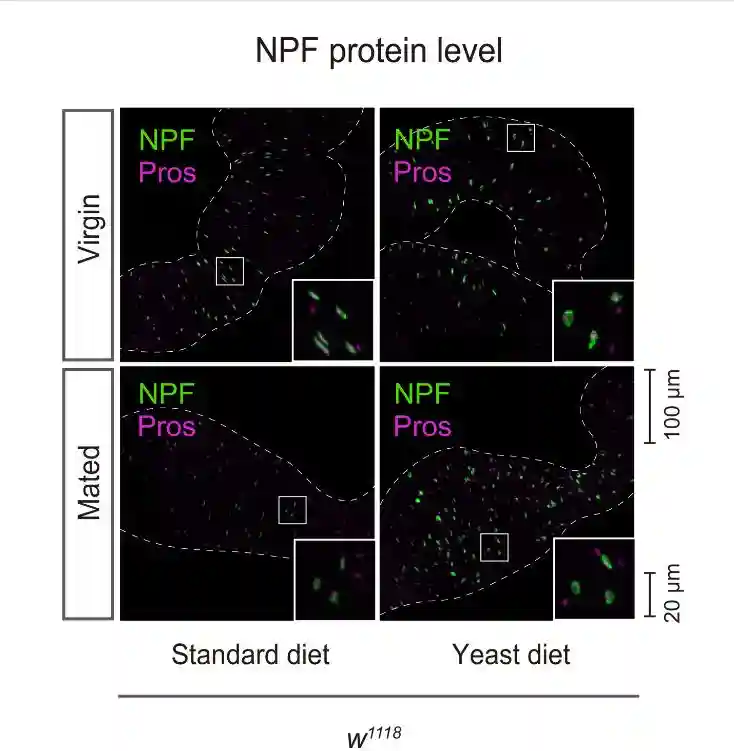
交配诱导的GSC增加需要NPF,NPF在交配时从中肠EEC释放,并由卵巢体细胞接收。假设酵母饮食对这种GSC增加的损害是由于抑制肠内分泌NPF的释放。首先检查标准和酵母饮食中野生型雌性和交配雌性苍蝇的中肠EEC中NPF蛋白的含量。
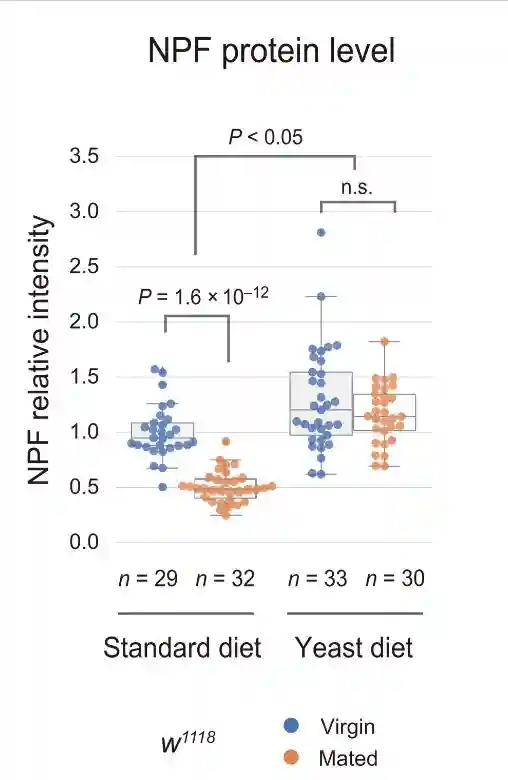
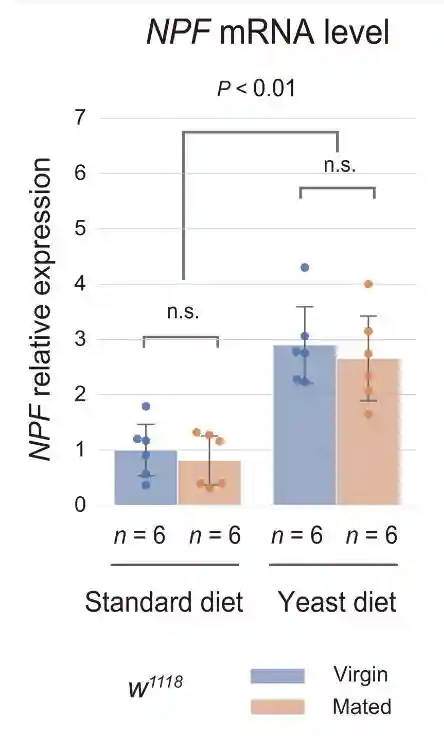
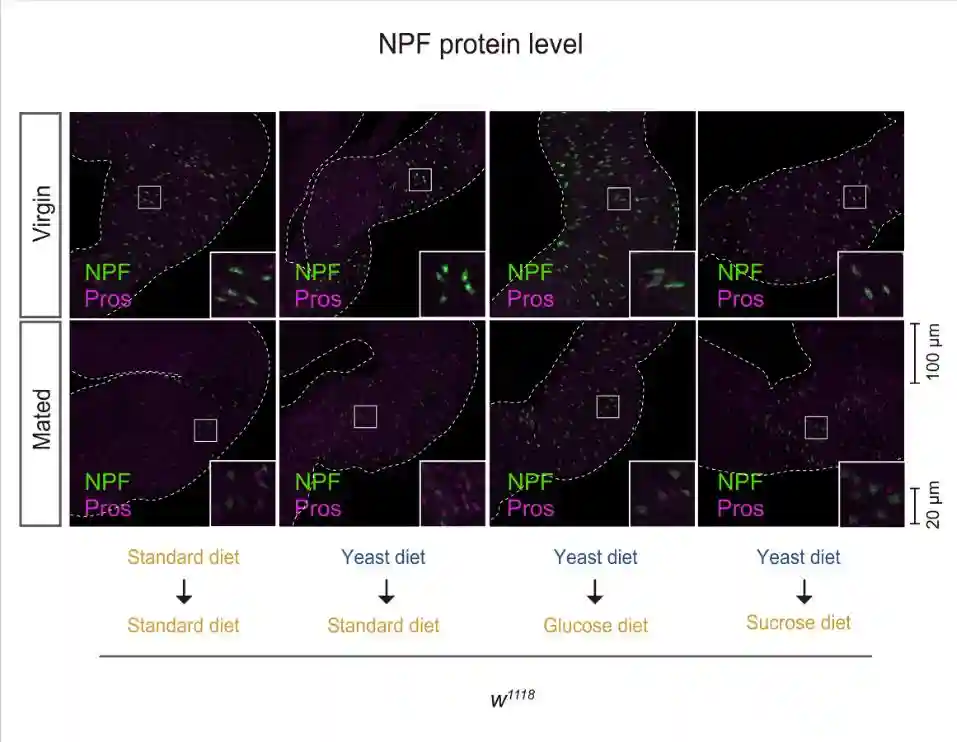
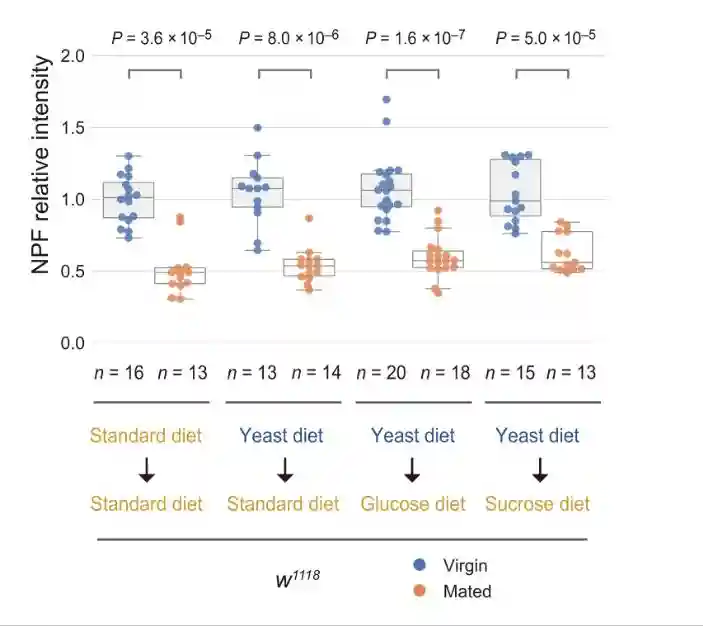
在标准饮食中,与雌性相比,交配雌性EEC中的肠内分泌NPF蛋白水平降低。然而,雌性和交配雌性之间肠道中的NPFmRNA丰度没有变化。NPF蛋白的高积累而没有NPFmRNA增加表明EECs的NPF分泌失败。
在酵母饮食中,雌性和交配雌性之间的NPF蛋白水平没有变化。在酵母饮食中,虽然雌性和交配雌性的NPFmRNA水平都增加,但交配并没有诱导肠道中NPFmRNA水平的增加。这些结果表明,即使在交配后,中肠EEC的NPF释放也被酵母饮食抑制。
通过使用玻璃针将合成NPF肽注射到成年雌性中来手动递送,与向雌性注射磷酸盐缓冲盐水相比,在标准饮食中向野生型雌性注射NPF导致GSC数量增加。将NPF注射到酵母饮食中的野生型雌性中也诱导了GSC数量的增加,无论它们是雄性还是交配的雌性。
NPF注射诱导的GSC增加与雌性和交配雌性GSC的pMad强度升高密切相关。TKg-GAL4介导的NPF在NPFEEC中的过表达并没有完全恢复交配诱导的GSC增加对酵母饮食的失败,表明NPF释放由酵母饮食控制。
结果表示交配后中肠EEC释放的NPF被酵母饮食抑制,导致通过Dpp生态位信号传导损害交配诱导的GSC增加。
酵母饮食和其他饮食之间的营养差异必须解释交配诱导的GSC增加的差异,这是由肠内分泌NPF介导的,EEC释放NPF受到雌性膳食糖的刺激。液相色谱-质谱分析显示,酵母日粮水解物中的葡萄糖含量低于标准日粮、葡萄糖、玉米面、YG和YC日粮中的葡萄糖。
用标准和酵母饮食饲养野生型苍蝇,将这些苍蝇转移到标准饮食,酵母饮食或含有蔗糖和琼脂的饮食,在转移后进行5天的培养期。允许其中一些苍蝇与雄配1天,其他雌性苍蝇在没有雄性的情况下连续饲养1天。
交配后,GSC在所有日粮中饲养的果蝇中增加,即使是那些事先用酵母日粮饲养的果蝇。考虑到葡萄糖和蔗糖饮食分别仅含有葡萄糖和蔗糖作为主要营养素,这些结果表明膳食糖对于交配诱导的GSC增加是必要且足够的。
野生型苍蝇的肠内分泌NPF蛋白水平,这些苍蝇最初喂食酵母日粮转移到标准,葡萄糖或蔗糖饮食。在酵母饮食后给予标准、葡萄糖或蔗糖饮食的苍蝇交配后,EEC中的NPF蛋白积累减少。这些结果表明,膳食糖对于交配诱导的EEC中释放的NPF也是必要且足够的。
雌性中肠EEC的糖反应性NPF释放需要低亲和力葡萄糖转运蛋白溶质载体家族2蛋白,糖转运蛋白1。Sut1对于交配诱导的NPF从EEC释放也必须是必需的。
在由TKg-GAL1驱动的Sut4RNA干扰的雌性中,其在NPFEEC中是活跃的。在标准饮食的苍蝇交配后,EEC中的NPF蛋白水平仍然降低。
在标准饮食中,在TKg-GAL4驱动的Sut1RNAi雌性中观察到交配诱导的GSC增加。实验表明,Sut1对交配后EEC释放的NPF没有主要贡献。
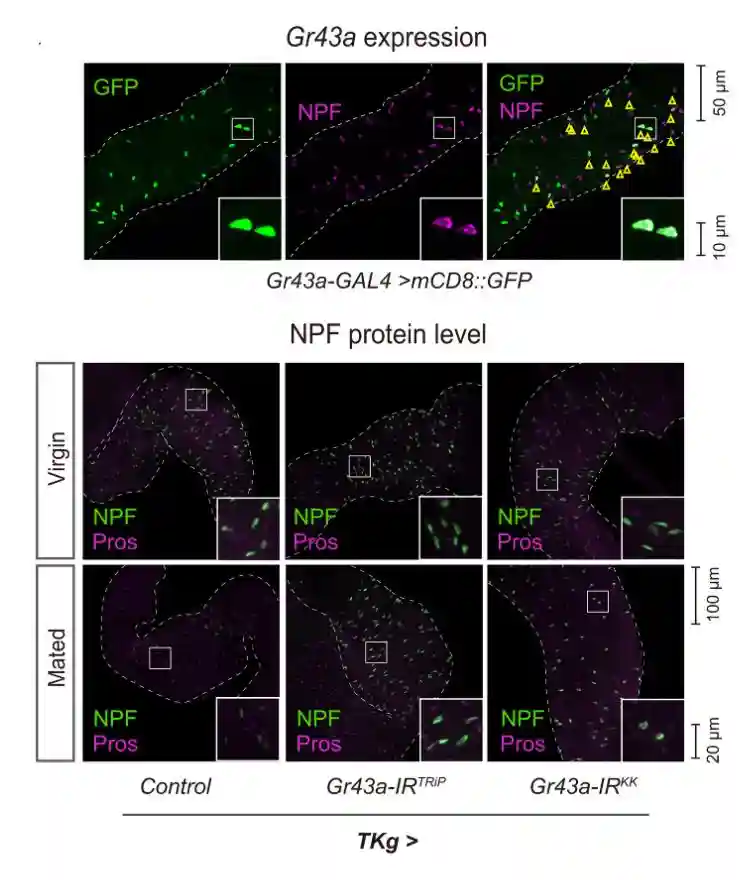
为了阐明交配后糖响应NPF释放的机制,专注于负责糖检测的Grs。在哺乳动物中,存在于EEC中的一些Grs接受营养并调节肠内分泌激素释放响应营养。在黑腹果蝇中,一些Gr基因在NPFEEC中表达,尽管它们在EEC中的功能尚未确定。进行转基因RNAi实验,以检查EEC中的哪种Gr参与交配诱导的GSC增加。
使用R13G46-GAL06敲低了4个Gr基因中的每一个,据报道在NPFEEC中具有活性。在13种GRS中,7种在EEC中表达,而9种已被表征为糖味受体。发现编码苦味受体的基因RNAi和一些糖味受体对交配诱导的GSC增加没有影响。
在编码糖味受体的Gr64a的RNAi的情况下,雌性的GSC数量异常高。编码三种糖味受体Gr5a,Gr43a和Gr64f的基因RNAi损害了交配诱导的GSC增加。Gr5a和Gr43a分别是用于感测海藻糖和果糖的味觉受体,而Gr64f可以感测多种糖,包括葡萄糖和蔗糖。
葡萄糖饮食中没有可检测到的果糖,葡萄糖饮食足以确保交配诱导的GSC增加。交配诱导的NPF释放需要果糖受体Gr43a的事实增加了葡萄糖衍生代谢物参与交配诱导的GSC增加的可能性。
交配前后以标准饮食饲养的野生型苍蝇的代谢组学变化,与交配雌性全身的代谢变化相比,血淋巴中的代谢物在它们之间发生了剧烈变化。
在183种代谢物中,43种血淋巴代谢物表现出0倍以上的变化。赤藓糖6-磷酸、果糖6-磷酸、5-磷酸葡萄糖酸、核糖、核酮糖7-磷酸和景天庚酮糖5-磷酸,所有这些都是通过戊糖磷酸途径产生的,与雌性相比,交配雌性升高。
交配后升高的其他血淋巴代谢物是糖,包括葡萄糖、山梨糖醇、海藻糖和果糖。这些结果支持了血淋巴果糖对于调节交配诱导的NPF分泌和GSC增加至关重要。
在真核生物中,果糖是通过多元醇途径从葡萄糖合成的,其中葡萄糖首先转化为山梨糖醇,然后转化为果糖。多元醇途径的关键酶之一是山梨醇脱氢酶,负责将山梨糖醇转化为果糖。
检查Sodh突变雌性和交配雌性的血淋巴糖水平,在这两种情况下,与雌性相比,交配雌性的血淋巴山梨糖醇急剧上调。这一观察结果符合Sodhs的生化功能,因为如果Sodh被消除,酶底物山梨糖醇应该会积累。
发现交配诱导的血淋巴果糖升高在Sodh突变雌性中受到抑制。表型不太可能是由于交配失败造成的,因为几乎所有对照和Sodh突变体的雌性都交配了。这些结果表明,多元醇途径是交配后血淋巴果糖升高所必需的。
一般来说,膳食糖,特别是葡萄糖,直接影响几乎所有生物体细胞的代谢状态,包括黑腹果蝇种系谱系细胞。膳食葡萄糖不太可能直接作用于GSC或其他卵巢细胞来调节交配诱导的GSC增加。
相反膳食葡萄糖有助于由称为多元醇途径的代谢途径产生的血淋巴果糖升高。交配诱导的肠内分泌NPF通过果糖特异性GrGr43a从中肠释放,然后是交配后NPF介导的GSC增加,需要果糖升高。
研究表明,膳食糖是交配后GSC增加不可或缺的先决条件,从而增强产蛋量。研究结果还表明,不仅蛋白质,而且糖都是影响GSCs的必需膳食营养素。